
Cyklina - Cyclin
Cyklina to rodzina białek, które kontrolują progresję komórki przez cykl komórkowy poprzez aktywację enzymów kinazy zależnej od cyklin (CDK) lub grupy enzymów niezbędnych do syntezy cyklu komórkowego .
Etymologia
Cykliny zostały pierwotnie odkryte przez R. Timothy'ego Hunta w 1982 roku podczas badania cyklu komórkowego jeżowców.
W wywiadzie dla „The Life Scientific” (wyemitowanym 13.12.2011) prowadzonym przez Jima Al-Khalili , R. Timothy Hunt wyjaśnił, że nazwa „cyklina” pochodzi od jego hobbystycznego kolarstwa. Dopiero po nazewnictwie ujawniło się jego znaczenie w cyklu komórkowym. Jak było na miejscu nazwa utknęła. R. Timothy Hunt : „Tak przy okazji, nazwa cyklina, którą ukułem, była naprawdę żartem, ponieważ tak bardzo lubiłem wtedy jeździć na rowerze, ale oni przychodzili i odchodzili w celi…”
Funkcjonować

Cykliny zostały pierwotnie nazwane, ponieważ ich stężenie zmienia się cyklicznie podczas cyklu komórkowego. (Zauważ, że cykliny są teraz klasyfikowane zgodnie z ich konserwatywną strukturą kasety cyklinowej i nie wszystkie te cykliny zmieniają swój poziom w cyklu komórkowym.) Oscylacje cyklin, a mianowicie fluktuacje ekspresji genu cykliny i niszczenie przez szlak proteasomów, w którym pośredniczy ubikwityna , indukują oscylacje aktywności Cdk w celu kierowania cyklem komórkowym. Cyklina tworzy kompleks z Cdk, który zaczyna się aktywować, ale pełna aktywacja wymaga również fosforylacji. Tworzenie kompleksu powoduje aktywację miejsca aktywnego Cdk . Same cykliny nie mają aktywności enzymatycznej, ale mają miejsca wiązania dla niektórych substratów i kierują Cdk do określonych lokalizacji subkomórkowych.
Cykliny po związaniu z kinazami zależnymi , takimi jak białko p34 / cdc2 / cdk1 , tworzą czynnik sprzyjający dojrzewaniu . MPF aktywują inne białka poprzez fosforylację . Te fosforylowane białka są z kolei odpowiedzialne za określone zdarzenia podczas podziału cyklu, takie jak tworzenie mikrotubul i przebudowa chromatyny . Cykliny można podzielić na cztery klasy na podstawie ich zachowania w cyklu komórkowym komórek somatycznych kręgowców i komórek drożdży: cykliny G1, cykliny G1/S, cykliny S i cykliny M. Podział ten jest przydatny, gdy mówimy o większości cykli komórkowych, ale nie jest uniwersalny, ponieważ niektóre cykliny mają różne funkcje lub czasy w różnych typach komórek.
Cykliny G1/S rosną w późnym G1 i spadają we wczesnej fazie S. Kompleks cykliny Cdk-G1/S zaczyna indukować początkowe procesy replikacji DNA, głównie poprzez zatrzymanie układów, które zapobiegają aktywności Cdk fazy S w G1. Cykliny promują również inne działania mające na celu postęp cyklu komórkowego, takie jak duplikacja centrosomu u kręgowców lub trzon wrzeciona u drożdży. Wzrostowi obecności cyklin G1/S towarzyszy wzrost cyklin S.
Cykliny G1 nie zachowują się jak inne cykliny, w tym sensie, że stężenia rosną stopniowo (bez oscylacji) przez cały cykl komórkowy w oparciu o wzrost komórki i zewnętrzne sygnały regulujące wzrost. Obecność cyklin G koordynuje wzrost komórek wraz z wejściem w nowy cykl komórkowy.
Cykliny S wiążą się z Cdk, a kompleks bezpośrednio indukuje replikację DNA. Poziomy cyklin S pozostają wysokie, nie tylko w fazie S, ale także poprzez G2 i wczesną mitozę, aby promować wczesne zdarzenia w mitozie.
Stężenie cykliny M wzrasta, gdy komórka zaczyna wchodzić w mitozę, a stężenie osiąga szczyt w metafazie. Zmiany komórkowe w cyklu komórkowym, takie jak montaż wrzecion mitotycznych i ustawienie siostrzanych chromatyd wzdłuż wrzecion, są indukowane przez kompleksy cyklina M-Cdk. Zniszczenie cyklin M podczas metafazy i anafazy, po spełnieniu Punktu Kontrolnego Montażu Wrzeciona, powoduje wyjście z mitozy i cytokinezy. Ekspresja cyklin wykrytych immunocytochemicznie w poszczególnych komórkach w odniesieniu do zawartości DNA komórkowego (faza cyklu komórkowego) lub w odniesieniu do inicjacji i zakończenia replikacji DNA podczas fazy S, może być mierzona za pomocą cytometrii przepływowej .
Herpeswirus mięsaka Kaposiego ( KSHV ) koduje cyklinę typu D (ORF72), która wiąże CDK6 i prawdopodobnie przyczynia się do nowotworów związanych z KSHV.
Struktura domeny
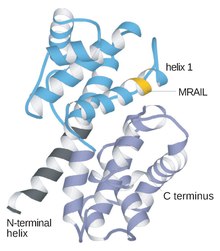
Cykliny na ogół bardzo różnią się od siebie strukturą pierwszorzędową lub sekwencją aminokwasową. Jednak wszyscy członkowie rodziny cyklin są podobni w 100 aminokwasach, które tworzą pudełko cyklin. Cykliny zawierają dwie domeny podobnego fałdu all-α , pierwsza zlokalizowana na N-końcu, a druga na C-końcu . Uważa się, że wszystkie cykliny zawierają podobną strukturę trzeciorzędową dwóch zwartych domen 5 α helis. Pierwszym z nich jest zachowane pudełko cyklin, poza którym cykliny są rozbieżne. Na przykład, regiony końca aminowego cyklin S i M zawierają krótkie motywy destrukcyjnej skrzynki, które nakierowują te białka na proteolizę w mitozie.



Rodzaje
Istnieje kilka różnych cyklin, które są aktywne w różnych częściach cyklu komórkowego i powodują, że Cdk fosforyluje różne substraty. Istnieje również kilka „sierocych” cyklin, dla których nie zidentyfikowano partnera Cdk. Na przykład, cyklina F jest cyklina sierocy, który jest niezbędny dla G 2 / M przejściowego. Badanie na C. elegans ujawniło specyficzne role cyklin mitotycznych. Warto zauważyć, że ostatnie badania wykazały, że cyklina A tworzy środowisko komórkowe, które sprzyja oderwaniu mikrotubul od kinetochorów w prometafazie, aby zapewnić skuteczną korekcję błędów i wierną segregację chromosomów. Komórki muszą precyzyjnie oddzielić swoje chromosomy, co polega na dwukierunkowym przyłączeniu chromosomów do mikrotubul wrzeciona poprzez wyspecjalizowane struktury zwane kinetochorami. We wczesnych fazach podziału istnieje wiele błędów w sposobie, w jaki kinetochory wiążą się z mikrotubulami wrzeciona. Niestabilne przyczepy sprzyjają korekcji błędów, powodując ciągłe odrywanie, przestawianie i ponowne dołączanie mikrotubul od kinetochorów w komórkach, gdy próbują znaleźć prawidłowe przyczepienie. Cyklina białkowa A zarządza tym procesem, utrzymując proces do momentu wyeliminowania błędów. W normalnych komórkach trwała ekspresja cykliny A zapobiega stabilizacji mikrotubul związanych z kinetochorami nawet w komórkach z wyrównanymi chromosomami. Wraz ze spadkiem poziomu cykliny A przyłączenia mikrotubul stają się stabilne, umożliwiając prawidłowy podział chromosomów w miarę postępu podziału komórki. Natomiast w komórkach z niedoborem cykliny A przyłączenia mikrotubul są przedwcześnie stabilizowane. W konsekwencji komórki te mogą nie korygować błędów, co prowadzi do wyższych wskaźników nieprawidłowej segregacji chromosomów.
Główne grupy
Istnieją dwie główne grupy cyklin:
- G 1 / S cykliny - niezbędne do regulacji cyklu komórkowego, w G 1 / S przejścia ,
- G 2 / Cykliny M - niezbędne do kontroli cyklu komórkowego w G2 / M przejścia ( mitozy ). G 2 cykliny / M gromadzić stale podczas G 2 i gwałtownie zniszczone wyjścia komórki z fazy mitozy (na końcu fazy M ).
Podtypy
Konkretne podtypy cyklin wraz z odpowiadającymi im CDK (w nawiasach) to:
| Gatunki | G1 | G1/S | S | M |
|---|---|---|---|---|
| S. cerevisiae | Cln3 (Cdk1) | Cln 1,2 (Cdk1) | Clb 5,6 (Cdk1) | Ćw 1,2,3,4 (Cdk 1) |
| S. pombe | Puc1? ( Cdc2 ) | Puc1, Cig1? (Cdc2) | Cig2, Cig1? (Cdc2) | Cdc13 (Cdc2) |
| D. melanogaster | cyklina D (Cdk4) | cyklina E (Cdk2) | cyklina E, A (Cdk2,1) | cyklina A, B, B3 (Cdk1) |
| X. laevis | albo nie wiadomo, albo nie występuje | cyklina E (Cdk2) | cyklina E, A (Cdk2,1) | cyklina A, B, B3 (Cdk1) |
| H. sapiens | cyklina D 1,2,3 ( Cdk4 , Cdk6 ) | cyklina E ( Cdk2 ) | cyklina A ( Cdk2 , Cdk1 ) | cyklina B ( Cdk1 ) |
| rodzina | członkowie |
|---|---|
| ZA | CCNA1 , CCNA2 |
| b | CCNB1 , CCNB2 , CCNB3 |
| do | CCNC |
| re | CCND1 , CCND2 , CCND3 |
| mi | CCNE1 , CCNE2 |
| fa | CCNF |
| sol | CCNG1 , CCNG2 |
| H | CCNH |
| ja | CCNI , CCNI2 |
| jot | CCNJ , CCNJL |
| K | CCNK |
| L | CCNL1 , CCNL2 |
| O | CCNO |
| P | CCNP |
| T | CCNT1 , CCNT2 |
| Tak | CCNY , CCNYL1 , CCNYL2 , CCNYL3 |
Inne białka zawierające tę domenę
Ponadto następujące białko ludzkie zawiera domenę cyklinową:
Historia
Leland H. Hartwell , R. Timothy Hunt i Paul M. Nurse zdobyli Nagrodę Nobla w 2001 r. w dziedzinie fizjologii lub medycyny za odkrycie cykliny i kinazy zależnej od cyklin.
Bibliografia
Dalsza lektura
- Krieger M, Scott MP, Matsudaira PT, Lodish HF, Darnell JE, Zipursky L, Kaiser C, Berk A (2004). Biologia komórki molekularnej (wyd. piąte). Nowy Jork: WH Freeman i CO. ISBN 0-7167-4366-3.
Linki zewnętrzne
- Eukariotyczny motyw liniowy, klasa motywu zasobów LIG_CYCLIN_1