
Radiodonta -Radiodonta
| Radiodonta Zakres czasowy: III stopień kambryjski – wczesny dewon ,
|
|
|---|---|

|
|
| Od lewej do prawej, od góry do dołu: Amplectobelua symbrachiata , Anomalocaris canadensis , Aegirocassis benmoulai , Peytoia nathorsti , Lyrarapax unguispinus , Cambroraster falcatus i Hurdia victoria | |
| Klasyfikacja naukowa | |
| Królestwo: | |
| Gromada: | |
| Klasa: |
Dinokaridida _
|
| Zamówienie: |
† Radiodonta
Collins , 1996 |
| Rodziny | |
|
|
Radiodonta to wymarły rząd stawonogów z grupy łodyg, który odniósł sukces na całym świecie w okresie kambryjskim . Mogą być określane jako radiodonty , radiodonty , radiodontids , anomalocarids lub anomalocaridids , chociaż dwa ostatnie pierwotnie odnoszą się do rodziny Anomalocarididae , która wcześniej obejmowała wszystkie gatunki tego rzędu, ale teraz jest ograniczona tylko do kilku gatunków. Radiodonty wyróżniają się charakterystycznymi wyrostkami czołowymi, które są zróżnicowane morfologicznie i wykorzystywane do różnych funkcji. Radiodonty obejmowały najwcześniejsze znane duże drapieżniki, ale także przesiewacze osadów i filtratory. Niektóre z najbardziej znanych gatunków radiodontów to taksony kambryjskie Anomalocaris canadensis , Hurdia victoria , Peytoia nathorsti , Titanokorys gainessii , Cambroraster falcatus i Amplectobelua symbrachiata , ordowicki Aegirocassis benmoulai i devonski Schinderhannes .
Etymologia
Nazwa Radiodonta (łac. promień „szprycha koła” i grecki odoús „ząb”) odnosi się do promieniowego układu płytek zębów (stożek ustny) otaczających usta, chociaż sugeruje się, że te cechy nie występują u niektórych gatunków radiodontów.
Definicja
Pierwotna diagnoza zakonu Radiodonta z 1996 roku przedstawia się następująco:
Radiodontids są dwustronnie symetrycznymi, wydłużonymi stawonogami z niezmineralizowanym naskórkiem, zazwyczaj najbardziej wytrzymałym w szczękach i pazurach. Ciało dzieli się na dwie tagmata , podobnie jak prosoma i opisthosoma stawonogów chelicerate . Zazwyczaj przednia część nie wykazuje zewnętrznej segmentacji, posiada jedną parę przedustnych pazurów, parę wyłupiastych oczu i brzuszną szczękę z promieniującymi zębami. Niektóre formy mają dodatkowe rzędy zębów i trzy lub cztery pary kończyn tylnych . Pień jest metameryczny , zwykle z około 13 segmentami bocznie rozwijającymi się tworzącymi się płatami do pływania i skrzela do oddychania i może kończyć się wydatnym, trzyczęściowym ogonem. Niektóre formy mają kończyny tułowia gnatobasic.
W 2014 r. klad Radiodonta został zdefiniowany filogenetycznie jako klad obejmujący wszelkie taksony bliższe Anomalocaris canadensis niż Paralithodes camtschaticus . W 2019 r. został przedefiniowany pod względem morfologicznym jako kompleks pancerza głowy zwierzęcia z elementami centralnymi (H-) i bocznymi (P-); wyrostki (endyty) z wyrostków czołowych z dodatkowymi kolcami; i zmniejszone przednie płatki lub pasma blaszek (łopatki łydkowe) i silne zwężenie ciała od przodu do tyłu.
Opis

Większość radiodontów była znacznie większa niż inne fauny kambryjskie , z typową długością ciała wahającą się od 30 do 50 centymetrów. Największym opisanym radiodontem jest ordowik Aegirocassis benmoulai , który mógł dorastać do dwóch metrów długości. Prawie kompletny okaz młodocianego Lyrarapax unguispinus mierzył zaledwie 18 milimetrów (0,71 cala), co czyni go jednym z najmniejszych znanych okazów radiodontów, chociaż dorosłe osobniki osiągały długość 8 centymetrów (3,1 cala). znana jest ponad połowa młodocianego Lyrarapax , ale nie wiadomo, czy ten okaz należy do osoby dorosłej. Największym znanym kambryjskim radiodontem był Laminacaris , chociaż znany tylko z przydatków czołowych, miał szacowaną długość ciała do 78,4 cm (30,9 cala) na podstawie Anomalocaris . Anomalocaris i Amplectobelua są również duże, osiągnęły 37,8 cm (14,9 cala) i 48 cm (19 cali) (według szacunków Houcaris saron (wcześniej Anomalocaris saron ) osiągnął 56 cm (22 cale), ale próbka użyta do oszacowania ciała długość nie należy już do tego gatunku); kambryjski hurdiid Titanokorys zbliżył się do niego pod względem wielkości, o szacowanej długości ciała około 50 centymetrów (20 cali).
Tułów radiodonta można podzielić na dwa rejony: głowę i tułów. Głowa składa się tylko z jednego segmentu ciała, znanego jako somit oczny, pokrytego sklerytami (kompleks pancerza głowy), posiada stawonogowe wyrostki czołowe, brzuszne narządy gębowe (stożek ustny) i szypułkowane oczy złożone . Zwężający się pień składa się z wielu segmentów ciała, z których każdy jest powiązany z parami klap i struktur podobnych do skrzeli (łopatki ostrogowe).
Wyrostek czołowy


Struktury przednie na głowie to para przydatków czołowych, które we wcześniejszych badaniach określano jako „pazury”, „przydatki chwytające”, „przydatki żerujące” lub „wielkie przydatki” (ostatni termin jest odradzany ze względu na homologię między wyrostki czołowe i oryginalne, megacheirańskie wielkie wyrostki są wątpliwe.). Są sklerotyzowane (utwardzone) i stawonogowe (segmentowane), posiadają brzuszne endyty (kolce) na większości swoich podomerów (jednostek segmentowych), a endyty mogą mieć dodatkowe rzędy pomocniczych kolców na ich przednich i tylnych krawędziach. Wyrostek czołowy składa się z dwóch obszarów: trzonu (w niektórych badaniach „szypułka”, „podstawa” lub „obszar bliższy”) oraz dystalnego regionu stawowego (nazywanego również „pazurem”). Po stronie brzusznej między podomerami może pojawić się trójkątny obszar pokryty miękkim naskórkiem (błona stawowa), który zapewnia elastyczność. Ich przedoczne i przedmózgowe pochodzenie sugeruje, że są one homologiczne z głównymi czułkami Onychophora i obrąbkiem Euarthropoda (wszystkie powstały z ocznego somitu ) , a nie homologiczne z chelicerae Chelicerata ani czułkami lub " wielkimi wyrostkami " innych stawonogów, które są deuto-mózgowe (powstały z postocznego somitu 1). Ponieważ morfologia wyrostków czołowych, zwłaszcza kolców, zawsze różni się między gatunkami, jest to jeden z najważniejszych sposobów identyfikacji gatunku. W rzeczywistości wiele radiodontów jest znanych tylko z kilku skamieniałych wyrostków czołowych.

Czołowe wyrostki Anomalocarididae , Amplectobeluidae i prawdopodobnie spokrewnione gatunki

Czołowe wyrostki Tamisiocarididae

Czołowe wyrostki Hurdiidae



Stożek ustny

Usta znajdują się po brzusznej stronie głowy, za punktem przyczepu przydatków czołowych i są otoczone pierścieniem płytek zębowych, tworzących część gębową znaną jako stożek ustny („szczęki” we wcześniejszych badaniach). 3 lub 4 płytki zębowe mogą być powiększone, nadając stożkowi jamy ustnej wygląd triradialny (np . Anomalocaris ) lub tetraradialny (np . Hurdiidae , Lyrarapax ). Wewnętrzny brzeg płytek ma kolce skierowane w stronę otworu gębowego. Dodatkowe rzędy wewnętrznych płytek zębów mogą występować w niektórych hurdiid rodzajów. Szczegółowa rekonstrukcja niektórych stożków amplectobeluid ustnych ma charakter spekulacyjny, ale prawdopodobnie nie przedstawiały one typowego układu promieniowego.
Skleryty głowy, oczu i tułowia

Kompleks trzech sklerytów głowy ( karapaks ) utworzony przez centralny element H (skleryt przedni lub tarcza głowy) i parę elementów P (skleryty boczne) pokrywają powierzchnię grzbietową i boczno-brzuszną głowy zwierzęcia. Elementy P mogą łączyć się ze sobą, jak również z elementem H, za pomocą wąskiego przedniego wydłużenia (szyja elementu P lub „dziób”). Skleryty głowy są małe i jajowate u Anomalocarididae i Amplectobeluidae , ale często powiększone u Hurdiidae , co odpowiada ich wyraźnym kształtom ciała (opływowe u Anomalocarididae / Amplectobeluidae, ale często zwarte u Hurdiidae). Głowa miała dwa szypułkowane oczy złożone , które mogły mieć ruchomość i znajdują się pomiędzy szczelinami utworzonymi przez tylne obszary elementu H i elementu P.


W przeciwieństwie do pierwotnej diagnozy, podział segmentów ciała (granice segmentów) może być widoczny z zewnątrz i żaden znany członek Radiodonta (z wyjątkiem domniemanego radiodonta Cucumericrus ) nie ma wyrostków tułowia (nogi). Tułów ma liczne segmenty tułowia ( somity ), zwężające się od przodu do tyłu, przy czym przednie 3 lub 4 segmenty są znacznie zwężone w okolicę szyi.

Odmiany płatów radiodontowych
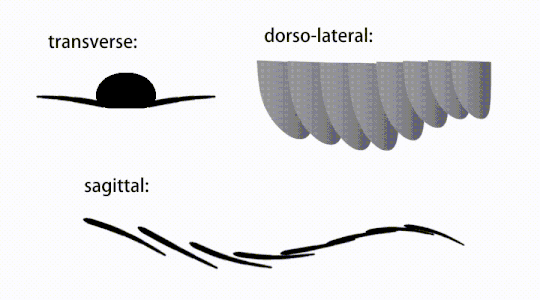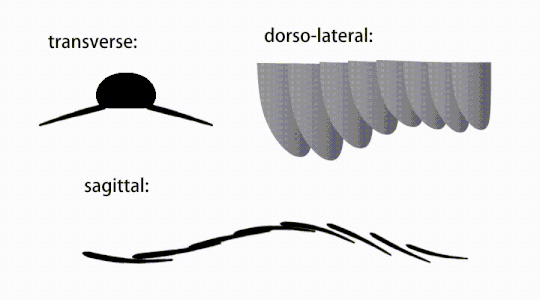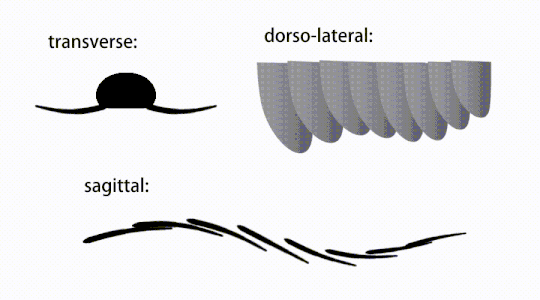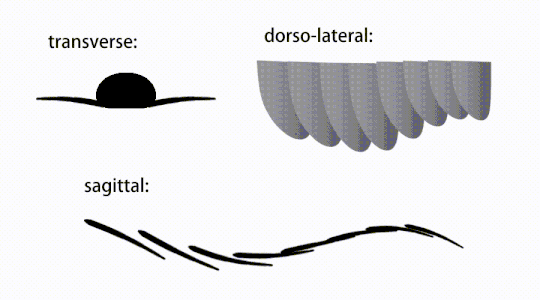
Ruch trzepoczących wyrostków radiodontów

Widok brzuszny uogólnionego radiodontu z GLS, ukazujący struktury przypominające gnathobase (GLS) związane ze zredukowanymi płatami przednimi



Przydatki tułowia były płatami podobnymi do płetw (w niektórych badaniach „płatami bocznymi” lub „płatami”), zwykle po jednej parze płatów brzusznych na segment ciała, z których każda nieznacznie zachodziła na jedną znajdującą się z przodu, ale dodatkowe, nienakładające się zestawy małych płatów grzbietowych może występować u niektórych gatunków Hurdiid . Płaty mogą mieć liczne struktury przypominające żyły (określane jako „promienie wzmacniające”, „promienie klapowe”, „pręty poprzeczne”, „linie poprzeczne” lub „żyły”). Klapy na szyi (nazywane „płatami zredukowanymi”, „płatami na szyję”, „płatami na głowę”, „płatami przednimi” lub „płatami zróżnicowanymi”) są znacznie zmniejszone. U niektórych gatunków, z każdej podstawy ich zredukowanych płatów szyjnych wyrastały przypominające szczękę przydatki pokarmowe zwane strukturami podobnymi do gnathobase (GLS). Liczne wydłużone wypustki przypominające ostrza (nazywane ostrzami lancetowatymi lub blaszkami) ułożone w rzędzie, tworzące pasma struktur podobnych do skrzeli , znanych jako ostrza osadzone, pokrywały grzbietową powierzchnię każdego segmentu ciała. Przynajmniej w Aegirocassis każde z lancetowatych ostrzy jest pokryte zmarszczkami. Płaty brzuszne mogą być homologiczne z endopodami dwuramiennych kończyn eustawonogów i lobpodów (lobopodów) skrzelowych lobopodów , a klapy grzbietowe i łopatki łokciowe mogą być homologiczne z wypustkami i blaszkami grzbietowymi zawierającymi skrzela dawnych taksonów. Pień może kończyć się wachlarzem ogonowym złożonym z 1 do 3 par łopatek, parą długich futer, wydłużoną strukturą końcową lub pozbawioną cech charakterystycznych tępą końcówką.

Stanleycaris , Hurdia , Aegirocassis , Peytoia i Cambroraster to przykłady radiodontów hurdiidowych.

Anomalocaris jest członkiem rodziny anomalocarididae , która w pewnym momencie obejmowała wszystkie radiodonty, ale obecnie obejmuje tylko kilka rodzajów.

Amplectobelua i Lyrarapax są przedstawicielami amplectobeluidae , która jest bardzo inkluzywną rodziną głównie chińskich radiodontów.

„Anomalocaris” briggsi i Tamisiocaris są przykładami rodziny tamisiocarididae , które były radiodontami żywiącymi się wyłącznie zawiesiną z kambru.



Struktury wewnętrzne

Ślady mięśni , układu pokarmowego i nerwowego opisano na podstawie niektórych skamieniałości radiodontów. Do płatów brzusznych zlokalizowanych w bocznych jamach każdego segmentu ciała połączono pary dobrze rozwiniętych mięśni. Pomiędzy mięśniami bocznymi znajduje się wyrafinowany układ trawienny, utworzony przez poszerzenie przedniego i tylnego jelita, oba połączone wąskim jelitem środkowym połączonym z sześcioma parami uchyłków jelitowych (gruczoły trawienne). W porównaniu z trójsegmentowymi mózgami eustawonogów i dwusegmentowymi mózgami onychoforanów , mózg radiodontów składa się tylko z jednego segmentu mózgu pochodzącego z somitu ocznego, protomózgowia. Nerwy przydatków czołowych i oczu złożonych powstały z przedniego i bocznego obszaru mózgu. Z tyłu mózgu znajdowała się para pozornie niepołączonych brzusznych sznurów nerwowych, które biegły przez obszar szyi zwierzęcia.
Paleoekologia
Fizjologia

Radiodonty zostały zinterpretowane jako zwierzęta nektoniczne lub nektobentosowe , a ich morfologia sugeruje aktywny styl życia. Muskularne, zachodzące na siebie klapy brzuszne mogły napędzać zwierzę przez wodę, prawdopodobnie poruszając się w formacji przypominającej falę, przypominającą współczesne płaszczki i mątwy . Pary klap grzbietowych, które u niektórych gatunków tworzą wachlarz ogonowy, mogły pomóc w sterowaniu i/lub stabilizacji zwierzęcia podczas lokomocji. W przypadku Anomalocaris morfologia wachlarza ogonowego sugeruje nawet, że może on szybko i skutecznie zmienić kierunek pływania. Z drugiej strony, niektóre hurdiidae mają cechy znacznie wyspecjalizowane w nektobentosowym stylu życia, takie jak Cambroraster z elementem H przypominającym kopułę, podobnym do pancerza kraba podkowy . Opaski nasadowych ostrzy z pomarszczonymi lancetowatymi ostrzami mogły zwiększać powierzchnię, co sugeruje, że były to skrzela , zapewniające zwierzęciu funkcję oddechową . Obfitość szczątków sklerityzowanych struktur, takich jak rozczłonkowane przydatki czołowe i kompleksy głowy sklerytu, sugeruje, że wśród radiodontów mogły występować masowe zjawiska linienia, co zostało również odnotowane u niektórych innych stawonogów kambryjskich , takich jak trylobity .
Dieta






Radiodonty miały różne strategie żywienia, które można było sklasyfikować jako drapieżniki drapieżne , przesiewacze osadów lub zawiesiny filtrujące . Na przykład drapieżne drapieżniki, takie jak Anomalocaris i Amplectobeluids , mogły być w stanie złapać zwinną zdobycz za pomocą drapieżnych wyrostków czołowych; ta ostatnia miała nawet mocny endit do trzymania zdobyczy jak szczypce . Przy mniejszym kompleksie pancerza głowy i dużej powierzchni błon stawowych, wyrostki czołowe tych taksonów miały większą elastyczność. Grube czołowe wyrostki przesiewaczy osadów, takich jak Hurdia i Peytoia , mają ząbkowane endyty z krzywizną mezjalną, które mogą tworzyć przypominającą koszyk pułapkę do grabienia osadów i przekazywania żywności w kierunku dobrze rozwiniętego stożka jamy ustnej. Endites przednich wyrostków z zawieszonych/filtrujących podajników, takich jak Tamisiocaris i Aegirocassis , mają elastyczne, gęsto upakowane kolce pomocnicze, które mogą odfiltrować składniki organiczne, takie jak mezozooplankton i fitoplankton , do 0,5 mm. Czołowe wyrostki Caryosyntrips , które są nietypowe dla radiodontów, ponieważ mają przeciwny kierunek powierzchni zawierających endyty i mogą być w stanie manipulować i miażdżyć zdobycz ruchem tnącym lub chwytającym podobnym do nożyc.
Stożki ustne radiodontów mogły być używane do odsysania i/lub gryzienia. Wraz z dużą różnorodnością wyrostków czołowych u różnych gatunków radiodontów, zróżnicowanie czopków jamy ustnej między gatunkami sugeruje również preferencje dotyczące różnych diet. Na przykład, trójpromieniowy stożek ustny Anomalocaris z nieregularnymi, gruźliczymi płytkami zębowymi i małym otworem mógł być przystosowany do małej i nektonicznej zdobyczy; podczas gdy sztywne tetraradialne stożki ustne Peytoia , Titanokorys , Hurdia i jeden izolowany szyszek ustny przypisywany Cambrorasterowi z większym otworem i czasami dodatkowymi płytkami zębów mogły być w stanie spożywać większe produkty żywnościowe w stosunku do ich wielkości ciała i prawdopodobnie bentosową lub endobentosową zdobycz .
Klasyfikacja
Powinowactwa taksonomiczne
|
|||||||||||||||||||||||||||||||||||||||||||||
| Podsumowanie filogenezy między Radiodonta a innymi taksonami Ecdysozoa |





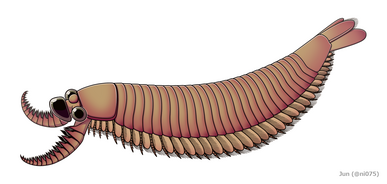
Radiodont/ opabiniid - pośrednia eustawonoga Kylinxia

Schematyczna rekonstrukcja bazalnego deuteropoda Erratusa


Większość analiz filogenetycznych sugeruje, że radiodonty, obok opabiniidów ( Opabinia i Utaurora ), należą do stawonogów z grupy łodyg tylko od podstawy do deuteropoda , kladu obejmującego górny pień (np . fuxianhuiidae i dwuskorupowe stawonogi) i Euarthropoda koronowe (np . Artiopoda , Mandibulicerata i Chelicerata ). Interpretacja ta jest poparta licznymi planami stawonogów znalezionymi na radiodontach i opabiniidach, takich jak szypułkowane oczy złożone , gruczoły trawienne, przydatki tułowia tworzące się przez elementy grzbietowe i brzuszne (prekursor stawonogów dwuramiennych). W porównaniu do opabiniidów, które mają tylne otwarcie ust i zrośnięte prawie przydatki czołowe (porównywalne do eustawonoga tylnego zespołu obrąbka /hipostomu), z drugiej strony radiodonty miały skleryt grzbietowy podobny do eustawonoga (element H) i stawonogów (przydatki czołowe). regiony głowy, wzdłuż łuskowatych końców jelit. Fakt, że zarówno radiodonty, jak i opabiniidae nie mają egzoszkieletu w okolicy tułowia, sugeruje, że pochodzenie oczu złożonych i stawonogów (segmentowanych przydatków) poprzedza artrodyzację (pełny zestaw egzoszkieletu tułowia) w linii pnia stawonogów. Zwężony obszar szyi z wyrostkami żywiącymi niektórych radiodontów może również rzucić światło na pochodzenie wyrafinowanej głowy stawonoga, która powstała w wyniku połączenia wielu przednich segmentów ciała. Podstawowe deuteropody, które posiadają mieszankę cech radiodontów/opabiniidów, takich jak Kylinxia i Erratus , mogą reprezentować formy pośrednie między radiodontami, opabiniidami i innymi eustawonogami.

Siberiid Lobopodians Siberion ( u góry po lewej), Megadictyon (na dole pośrodku) i Jianshanopodia (u góry po prawej)
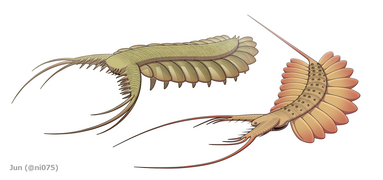
Skrzelowe lobopody Pambdelurion ( po lewej) i Kerygmachela (po prawej)

Megacheira lub „wielkie stawonogi wyrostka robaczkowego”, klasa możliwych chelicerate pnia , wcześniej uważana za bliską krewną radiodonta.



Taksony po prostu podstawowe dla gałęzi radiodonta, opabiniidae i euarthropod są " skrzelowymi lobopodianami " takimi jak Pambdelurion i Kerygmachela , które czasami łączyły się pod klasą Dinocaridida z opibiidami i radiodontami. Mają płaty ciała, gruczoły trawienne, duże (prawdopodobnie złożone) oczy i wyspecjalizowane wyrostki czołowe, takie jak dawne taksony, ale ich wyrostki czołowe nie są ani stawonogów, ani zrośniętych, oczy nie są nieruchome, struktury podobne do skrzeli są mniej widoczne i z pewnością mają lobpody pod każdym z nich. ich klapy. Taksony, nawet podstawowe dla „skrzelonych lobopodów”, to syberydy, takie jak Megadictyon i Jianshanopodia , grupa lobopodów z potężnymi wyrostkami czołowymi i gruczołami trawiennymi, ale bez płatów ciała. Takie formy pośrednie między lobopodami a radiodontami/eustawonogami sugerują, że stawonogi należące do grupy całkowitej powstały z parafiletycznego gatunku lobopodów, wraz z dwoma pozostałymi istniejącymi typami panartropoda Tardigrada i Onychophora .
Wcześniejsze badania mogą sugerować radiodonty jako grupę inną niż stawonogi łodygowe, taką jak dotychczas nieznana gromada ; cycloneuralia n robaki uległy zbieżności ze stawonogami (w oparciu o promieniste narządy gębowe podobne do cykloneuralów); łodygi chelicerate euarthropod obok megacheira ns, znanych również jako stawonogi wielkich przydatków (w oparciu o podobieństwo między przydatkami czołowymi radiodonta, wielkimi przydatkami megacheira i chelicerae ); lub Schinderhannes bartelsi , który w ostatnich analizach został uznany za hurdiidowego radiodonta, jako gatunek bliżej spokrewniony z eustawonogami niż z innymi radiodontami (w oparciu o pewne domniemane cechy przypominające eustawonogi występujące u Schinderhannesa ). Jednak żaden z nich nie znalazł poparcia w późniejszych śledztwach. Promieniste narządy gębowe nie są wyłączne dla cykloneuralnych i są bardziej prawdopodobne jako rezultat konwergentnej ewolucji lub plesimorfii ekdyzozoa n , ponieważ znaleziono je również u panartropodów , takich jak niesporczaki i niektóre lobopodia ; radiodonty pozbawione cech charakterystycznych dla stawonogów, takich jak tergity tułowia i liczne przydatki głowy, a wielkie przydatki megacheira uważano za deutomózgowe, co może być niehomologiczne z przedmózgowymi przydatkami przedmózgowymi; domniemane cechy eustawonoga znalezione na pojedynczej skamieniałości Schinderhannesa są wątpliwe i mogą przedstawiać inne struktury podobne do radiodontów.
Powiązania
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Filogeneza Radiodonta według Moysiuka i Carona 2021 |





Tradycyjnie wszystkie gatunki radiodontów były umieszczane w jednej rodzinie, Anomalocarididae , stąd poprzednia nazwa zwyczajowa „anomalocaridid” i nadal była czasami używana w odniesieniu do całego rzędu, nawet po przeklasyfikowaniu. Od czasu zmiany przydziału dokonanej przez Vinther et al. W 2014 r. większość gatunków radiodontów została przeklasyfikowana w ramach trzech nowych rodzin: Amplectobeluidae , Tamisiocarididae (dawniej Cetiocaridae ) i Hurdiidae . Włączając Anomalocarididae, cztery ostatnie rodziny radiodontów mogą tworzyć klad Anomalocarida .
Pierwotny opis rzędu Radiodonta obejmował Anomalocaris , Laggania (później znany jako Peytoia ), Hurdia , Proboscicaris , Amplectobelua , Cucumericrus i Parapeytoia . Jednak Proboscicaris jest obecnie uważany za młodszy synonim Hurdia , a Parapeytoia jest uważana za Megacheirana . Ze względu na ograniczone odkrycie, pozycja Cucumericrus w obrębie Radiodonta jest niejasna, ponieważ albo nie została wyselekcjonowana przez analizę filogenetyczną , albo została rozwiązana w politomii za pomocą Radiodonta i Euarthropoda .
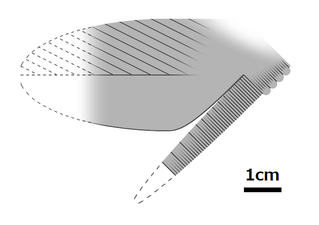
Jedna ze słabo poznanych części ciała (wyrostek tułowia) Cucumericrus decoratus ; gatunek ten może nie reprezentować prawdziwego radiodonta.

Wyrostek czołowy "Anomalocaris" briggsi . Pomimo sugerowanej nazwy rodzajowej, na podstawie analizy filogenetycznej jest mało prawdopodobne, aby ten radiodont był gatunkiem rodzaju Anomalocaris .

Pierwszą dogłębną analizę filogenetyczną Radiodonta przeprowadzili Vinther et al . w 2014 r., po którym nastąpiło kilka badań podrzędnych z mniej lub bardziej zmodyfikowanymi wynikami. W większości analiz Caryosyntrips jest najbardziej podstawowym rodzajem, ale został rozwiązany w politomii z innymi radiodontami i Euarthropoda (obok Cucumericrus , jeśli uwzględniono). Z wyłączeniem Caryosyntrips i Cucumericrus , powszechnie popiera się monofilię Radiodonta, a kilka wyników sugeruje możliwą parafilię (klad Anomalocarididae + Amplectobeluidae lub siostra Hurdiidae Euarthropoda). Domniemane synapomorfie monofiletycznej radiodonty obejmujące trójdzielny kompleks sklerytowy głowy i zróżnicowany obszar szyi. Rodzaj Anomalocaris w szerszym sensie zawsze uznawano za polifiletyczny , zwykle z „Anomalocaris” kunmingensis i „Anomalocaris” briggssi uznawanym za członka odpowiednio Amplectobeluidae i Tamisiocarididae . Wzajemne powiązania między Amplectobeluidae są niepewne, ponieważ powinowactwa do amplectobeluidów Lyrarapax i Ramskoeldia były czasami kwestionowane. Monofilia z rodziny gatunkowych Hurdiidae została odkryta w większości analiz i dobrze poparta kilkoma synapomorfiami (np. dystalny, przegubowy obszar wyrostka czołowego z bliższymi 5 podomerami z nierównymi końcami), przy czym często sugeruje się, że Tamisiocarididae są jej siostrzaną grupą.
-
Radiodonta
- ? Cucumericrus (domniemany radiodont)
- Caryosyntrips
-
Anomalokaryda
- Paranomalocaris (umieszczony w obrębie Anomalocarididae przez niektóre badania).
- Laminacaris (w niektórych badaniach umieszczany w Amlectobeluidae).
- Houcaris (umieszczony w obrębie Anomalocarididae, Amplectobeluidae lub Tamisiocarididae).
- Innowacje
-
Anomalocarididae
- Anomalocaris (w szerszym znaczeniu, niektóre gatunki można umieścić w innych rodzinach).
- Lenisicaris
-
Amplectobeluidae
- Lyrarapax (pozycja kwestionowana przez niektóre badania).
- Amplectobelua
- Ramskoeldia (stanowisko kwestionowane przez niektóre badania.)
- „Anomalocaris” kunmingensis (gatunki inne niż Anomalocaris czekają na zmianę nazwy. Oznaczone jako Amplectobelua kunmingensis przez Vinther et al. 2014.)
-
Tamisiocarididae
- Tamisiokaris
- „Anomalocaris” briggsi (gatunki inne niż Anomalocaris czekają na zmianę nazwy).
-
Hurdiidae
- Peytoia
- Schinderhannes
- Hurdia
- Aegirocassis
- Stanleycaris
- Pahwantia
- Urszulinakaris
- Kambroster
- ? Zhenghecaris (domniemany radiodont)
- Cordaticaris
- Bukaspina
- Tytanokorys
| Gatunek | Oryginalny opis | Nazwany rok | Rodzina | Wiek | Lokalizacja | Wyrostek czołowy | Kompleks sklerytów głowy |
|---|---|---|---|---|---|---|---|
| Cucumericrus decoratus | Hou, Bergström i Ahlberg | 1995 | (nieprzypisany) | Kambryjski etap 3 |
|
Nieznany | Nieznany |
| Caryosyntrips serratus | Daley i Budd | 2010 | (nieprzypisany) | Wuliuan – Drumian |
|

|
Nieznany |
| Caryosyntrips camurus | Pasztety i Daley | 2017 | (nieprzypisany) | Wuliuan |
|

|
Niekompletny |
| Caryosyntrips durus | Pasztety i Daley | 2017 | (nieprzypisany) | Drumian |
|

|
Nieznany |
| Paranomalocaris multisegmentalis | Wang, Huang i Hu | 2013 | Anomalocarididae ? | Kambryjski etap 4 |
|

|
Nieznany |
| Paranomalocaris simplex | Jiao, Pates, Lerosey-Aubril, Ortega-Hernandez, Yang, Lan, Zhang | 2021 | Anomalocarididae ? | Kambryjski etap 4 |
|

|
Nieznany |
| Chimera Laminacaris | Guo, Pates, Cong, Daley, Edgecombe, Chen i Hou | 2018 | (kontrowersyjny) | Kambryjski etap 3 |
|

|
Nieznany |
| Innovatiocaris maotianhanensis | Zeng, Zhao, Zhu | 2022 | (nieprzypisany) | Kambryjski etap 3 |
|

|
Niekompletny |
| Innowacje ? wielokolcowy | Zeng, Zhao, Zhu | 2022 | (nieprzypisany) | Kambryjski etap 3 |
|

|
Nieznany |
| Anomalocaris canadensis | Bielawy | 1892 | Anomalocarididae | Wuliuan |
|

|

|
| Lenisicaris pennsylvanica (dawniej Anomalocaris pennsylvanica ) | Reser | 1929 | Anomalocarididae | Kambryjski etap 3 |
|

|
Nieznany |
| Lenisicaris lupata | Wu, Ma, Lin, Sun, Zhang i Fu | 2021 | Anomalocarididae | Kambryjski etap 3 |
|

|
Nieznany |
| „Anomalocaris” kunmingensis | Wang, Huang i Hu | 2013 | Amplectobeluidae | Kambryjski etap 4 |
|

|
Nieznany |
| Houcaris magnabasis (dawniej Anomalocaris magnabasis ) | Pates, Daley, Edgecombe, Cong i Lieberman | 2019 | (kontrowersyjny) | Kambryjski etap 4 |
|

|
Nieznany |
| Houcaris saron (dawniej Anomalocaris saron ) | Hou, Bergström i Ahlberg | 1995 | (kontrowersyjny) | Kambryjski etap 3 |
|

|
Nieznany |
| „Anomalocaris” briggsi | Nedin | 1995 | Tamisiocarididae | Kambryjski etap 4 |
|

|
Nieznany |
| Ramskoeldia platyacantha | Cong, Edgecombe, Daley, Guo, Pasztety i Hou | 2018 | Amplectobeluidae | Kambryjski etap 3 |
|

|
Niekompletny |
| Ramskoeldia consimilis | Cong, Edgecombe, Daley, Guo, Pasztety i Hou | 2018 | Amplectobeluidae | Kambryjski etap 3 |
|

|
Niekompletny |
| Lyrarapax unguispinus | Cong, Ma, Hou, Edgecombe i Strausfield | 2014 | Amplectobeluidae | Kambryjski etap 3 |
|

|
|
| Lyrarapax trilobus | Cong, Daley, Edgecombe, Hou i Chen | 2016 | Amplectobeluidae | Kambryjski etap 3 |
|

|
|
| Amplectobelua symbrachiata | Hou, Bergström i Ahlberg | 1995 | Amplectobeluidae | Kambryjski etap 3 |
|

|

|
| Amplectobelua stephenensis | Daley i Budd | 2010 | Amplectobeluidae | Wuliuan |
|

|
Nieznany |
| Tamisiocaris borealis | Daley & Peel | 2010 | Tamisiocarididae | Kambryjski etap 3 |
|

|
Niekompletny |
| Ursulinacaris grallae | Pasztety, Daley i Butterfield | 2019 | Hurdiidae | Wuliuan |
|

|
Nieznany |
| Schinderhannes bartelsi | Kühl, Briggs i Rust | 2009 | Hurdiidae | Emsian |
|
Niekompletny | Niekompletny |
| Stanleycaris hirpex | Pates, Daley i Ortega-Hernández | 2018 | Hurdiidae | Wuliuan |
|

|
Pierwiastek P jest nieznany, prawdopodobnie nieobecny |
| Peytoia nathorsti | Walcott | 1911 | Hurdiidae | Wuliuan – Drumian |
|

|
Niekompletny |
| Peytoia infercambriensis (dawniej Cassubia infercambriensis ) | Lendzión | 1975 | Hurdiidae | Kambryjski etap 3 |
|

|
Nieznany |
| Aegirocassis benmoulai | Van Roy, Daley i Briggs | 2015 | Hurdiidae | Tremadocian |
|

|

|
| Hurdia wiktoria | Walcott | 1912 | Hurdiidae | Wuliuan – Drumian |
|

|

|
| Hurdia triangulata | Walcott | 1912 | Hurdiidae | Wuliuan |
|
|

|
| Cambroraster falcatus | Moysiuk i Caron | 2019 | Hurdiidae | Wuliuan |
|

|

|
| Pahvantia hasta | Robison i Richards | 1981 | Hurdiidae | Drumian |
|

|

|
| Cordaticaris striatus | Słońce, Zeng i Zhao | 2020 | Hurdiidae | Drumian |
|
Niekompletny |

|
| Zhenghecaris shankouensis | Vanner, Chen, Huang, Charbonnier i Wang | 2006 | Hurdiidae | Kambryjski etap 3 |
|
Nieznany |
 |
| Buccaspinea cooperi | Pates, Lerosey-Aubril, Daley, Kier, Bonino i Ortega-Hernández | 2021 | Hurdiidae | Drumian |
|

|
Nieznany |
| Wzmocnienie tytanokorysu | Caron i Moysiuk | 2021 | Hurdiidae | Wuliuan |
|

|

|
Historia


Wyrostek czołowy Anomalocaris canadensis

Stożek ustny Peytoia nathorsti

Element H z Hurdia victoria
![Sparowane wyrostki czołowe z nienazwanego radiodonta hurdiid[36]](https://upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg)
Sparowane wyrostki czołowe z nienazwanego radiodonta hurdiida



Historia radiodontów jest złożona. Niekompletne okazy należące do różnych części ciała tego samego gatunku były historycznie interpretowane jako należące do różnych gatunków, a nawet różnych typów. Przed uznaniem za grupę, okazy radiodontów przypisano do pięciu różnych typów: Porifera , Cnidaria , Echinodermata , Annelida i Arthropoda .
Pierwsze znane okazy radiodontów zostały zebrane ze złóż trylobitów na Mount Stephen przez Richarda G. McConnella z Geological Survey of Canada w 1886 lub 1888 roku. Okazy te zostały nazwane Anomalocaris canadensis w 1892 roku przez paleontologa GSC Josepha Whiteavesa . Whiteaves zinterpretował okazy, znane obecnie jako izolowane wyrostki czołowe, jako brzuch skorupiaka filokarowego . Dodatkowe okazy radiodontów zostały opisane w 1911 przez Charlesa Walcotta . Zinterpretował wyizolowany stożek ustny, który nazwał Peytoia nathorsti , jako meduzę, a słabo zachowany, ale stosunkowo kompletny okaz, który nazwał Laggania cambria , jako holoturian. W 1912 Walcott nazwał Hurdia victoria i H. triangulata na podstawie izolowanych pierwiastków H, które zinterpretował jako pancerze skorupiaków. Izolowane przednie wyrostki Peytoia i Hurdia , wspólnie znane jako „Wyrostek F” w Briggs 1979, zostały zidentyfikowane jako wyrostki Sidneyia w tym czasie. Element P Hurdia został nazwany Proboscicaris w 1962 roku i został zinterpretowany jako pancerz dwuskorupowego stawonoga.
Geological Survey of Canada zainicjował rewizję skamieniałości z łupków z Burgess w 1966 roku, nadzorowaną przez paleontologa z Cambridge University , Harry'ego B. Whittingtona . Ta rewizja ostatecznie doprowadziłaby do odkrycia kompletnego planu ciała radiodonta. W 1978 r. Simon Conway Morris uznał, że narządy gębowe Laggania są podobne do Peytoia , ale zinterpretował to jako dowód, że jest to złożona skamielina składająca się z meduzy Peytoia i gąbki. W 1979 roku Derek Briggs rozpoznał, że skamieliny Anomalocaris są wyrostkami, a nie brzuchami, ale zinterpretował je jako chodzące nogi obok „wyrostka F”. Dopiero w 1985 roku rozpoznano prawdziwą naturę skamieniałości Anomalocaris , Laggania i Peytoia i wszystkie zostały przypisane do jednego rodzaju, Anomalocaris . Następnie uznano, że Anomalocaris był formą odrębną od pozostałych dwóch, co spowodowało podział na dwa rodzaje, z których ten ostatni był różnie nazywany Laggania i Peytoia , dopóki nie ustalono, że oba reprezentują ten sam gatunek, a Peytoia ma pierwszeństwo. Później uznano, że część skamieniałości przypisanych do tych taksonów należała do innej formy, która została uznana za noszącą pancerz złożony z elementów Hurdia i Proboscicaris . Ostatecznie w 2009 roku okazy te zostały ponownie opisane jako Hurdia . Nawet po tych rozpoznaniach, częściowe błędne identyfikacje (np. stożek ustny i przednie wyrostki Peytoia zostały przypisane odpowiednio do Anomalocaris i Hurdia ) zostały również ujawnione w kolejnych badaniach.
Sam takson Radiodonta został ukuty w 1996 roku przez Desmonda Collinsa, po tym jak ustalono, że Anomalocaris i jego krewniacy reprezentują odrębną linię z powinowactwem do stawonogów, a nie nieznany dotąd typ. Collins ustanowił również klasę Dinocarida , aby zawierała rząd Radiodonta oraz Opabinidae , które rozpoznał jako odrębne ze względu na brak charakterystycznej struktury stożka jamy ustnej, jak radiodonty. Radiodonta po raz pierwszy otrzymała definicję filogenetyczną w 2014 roku. Pierwotnie Radiodonta była postrzegana jako zawierająca jedną rodzinę, Anomalocarididae , ale została podzielona na cztery rodziny w 2014 roku: Amplectobeluidae , Anomalocarididae, Cetiocaridae i Hurdiidae . Nazwa Cetiocaridae nie była zgodna z Międzynarodowym Kodeksem Nomenklatury Zoologicznej, dlatego w 2019 roku została przemianowana na Tamisiocarididae .
Do 2010 roku radiodonty były zazwyczaj uważane za jednolicie duże drapieżniki wierzchołkowe, ale odkrycia nowych gatunków w ciągu tej dekady doprowadziły do znacznego wzrostu znanej różnorodności ekologicznej i morfologicznej grupy.