
Wyższy zwój szyjki macicy - Superior cervical ganglion
| Górny zwój szyjki macicy (SCG) | |
|---|---|
 Schemat współczulnego szyjki macicy. (Oznaczony jako „Górny zwój szyjny”)
| |
| Detale | |
| Identyfikatory | |
| łacina | zwoju szyjki macicy superius |
| Siatka | D017783 |
| TA98 | A14.3.01.009 |
| TA2 | 6608 |
| FMA | 6467 |
| Anatomiczne terminy neuroanatomii | |
Lepsze zwoju szyjnego ( SCG ) stanowi część autonomicznego układu nerwowego (ANS), a dokładniej jest to część współczulnego układu nerwowego , oddział ANS najczęściej związanym z walki lub ucieczki . AUN składa się ze szlaków prowadzących do i ze zwojów , grup komórek nerwowych. Zwój pozwala na dużą rozbieżność w ścieżce neuronalnej, a także umożliwia bardziej zlokalizowane obwody do kontroli unerwionych celów. SCG to jedyny zwój we współczulnym układzie nerwowym, który unerwia głowę i szyję. Jest to największy i najbardziej rostralny (najwyższy) z trzech zwojów szyjnych . SCG unerwia wiele narządów, gruczołów i części układu szyjnego w głowie.
Struktura
Lokalizacja
SCG znajduje się naprzeciwko drugiego i trzeciego kręgu szyjnego . Leży głęboko do pochewki tętnicy szyjnej wewnętrznej i żyły szyjnej wewnętrznej oraz przed mięśniem Longus capitis . SCG zawiera neurony, które dostarczają współczulnego unerwienia do wielu narządów docelowych w obrębie głowy.
SCG przyczynia się również do powstania splotu szyjnego . Splot szyjny powstaje z połączenia przednich podziałów czterech górnych nerwów szyjnych. Każdy otrzymuje szary ramus communicans z górnego zwoju szyjnego pnia współczulnego.
Morfologia i fizjologia i jej neurony
Górny zwój szyjny ma kolor czerwono-szary i zwykle ma kształt wrzeciona ze zwężającymi się końcami. Czasami SCG jest szeroki i spłaszczony, a czasami zwężony w pewnych odstępach. Powstaje z połączenia czterech zwojów , odpowiadających czterem górnym nerwom szyjnym , C1-C4. Ciała tych przedzwojowych neuronów współczulnych są specyficznie zlokalizowane w rogu bocznym rdzenia kręgowego. Te neurony przedzwojowe następnie wchodzą do SCG i łączą się z neuronami postganglionowymi, które opuszczają rostralny koniec SCG i unerwiają narządy docelowe głowy.
Istnieje wiele typów neuronów w SCG, od neuronów o niskim progu do neuronów o wysokim progu. Neurony o niskim progu mają większą szybkość odpalania potencjału czynnościowego , podczas gdy neurony o wysokim progu mają niską szybkość odpalania. Kolejne rozróżnienie pomiędzy typami neuronów SCG dokonuje się poprzez immunobarwienie . Immunobarwienie umożliwia klasyfikację neuronów SCG jako dodatnich lub ujemnych dla neuropeptydu Y (NPY), który znajduje się w podgrupie neuronów wysokoprogowych. Niskoprogowe neurony NPY-ujemne to neurony sekretomotoryczne, unerwiające gruczoły ślinowe. Neurony o wysokim progu NPY-ujemne to neurony naczynioruchowe, unerwiające naczynia krwionośne. Neurony NPY-dodatnie o wysokim progu to neurony zwężające naczynia, które unerwiają tęczówkę i szyszynkę.
Unerwienie
SCG otrzymuje informacje z ośrodka rzęskowo - rdzeniowego . Centrum rzęskowo-rdzeniowe znajduje się pomiędzy regionami C8 i T1 rdzenia kręgowego w obrębie międzyprzyśrodkowej kolumny. Włókna przedzwojowe unerwiające SCG to nerwy rdzeniowe w odcinku piersiowym, które rozciągają się od obszaru T1-T8 centrum rzęskowo-rdzeniowego. Nerwy te wchodzą do SCG przez nerw współczulny szyjny. Dojrzały akson przedzwojowy może unerwić od 50 do 200 komórek SCG. Włókna zazwojowe następnie opuszczają SCG przez nerw szyjny wewnętrzny i nerw szyjny zewnętrzny. Ta ścieżka unerwienia SCG jest pokazana poprzez stymulację szyjnego nerwu współczulnego, który wywołuje potencjały czynnościowe zarówno w zewnętrznych, jak i wewnętrznych nerwach szyjnych. Te włókna postganglionowe przechodzą z unerwienia wieloaksonowego ich obiektów docelowych do mniej głębokiego unerwienia wieloaksonowego lub unerwienia pojedynczego aksonowego, gdy neurony SCG dojrzewają podczas rozwoju poporodowego.
Funkcjonować
Współczulny układ nerwowy
SCG zapewnia współczulne unerwienie struktur w obrębie głowy, w tym szyszynki , naczyń krwionośnych w mięśniach czaszkowych i mózgu, splotu naczyniówkowego , oczu, gruczołów łzowych, korpusu szyjnego , gruczołów ślinowych i tarczycy .
Szyszynka
Aksony zazwojowe SCG unerwiają szyszynkę i biorą udział w rytmie dobowym . Połączenie to reguluje produkcję hormonu melatoniny, który reguluje cykle snu i czuwania, jednak wpływ unerwienia neuronów SCG szyszynki nie jest do końca poznany.
Ciało szyjne
Aksony zazwojowe SCG unerwiają tętnicę szyjną wewnętrzną i tworzą splot szyjny wewnętrzny . Wewnętrzny splot szyjny przenosi pozazwojowe aksony SCG do oka, gruczołu łzowego , błon śluzowych jamy ustnej, nosa i gardła oraz licznych naczyń krwionośnych w głowie.
Oko
Aksony zazwojowe zwoju szyjnego górnego unerwiają oko i gruczoł łzowy i powodują zwężenie tęczówki i twardówki, rozszerzenie źrenic, poszerzenie szpary powiekowej i zmniejszenie produkcji łez. Te reakcje są ważne podczas reakcji ANS typu „ walcz lub uciekaj ”. Rozszerzenie źrenic pozwala na zwiększenie jasności widzenia, a zahamowanie gruczołu łzowego zatrzymuje produkcję łez, pozwalając na nieuszkodzone widzenie i przekierowanie energii w inne miejsce.
Naczynia krwionośne skóry
Aksony pozazwojowe SCG unerwiają naczynia krwionośne w skórze i powodują ich zwężenie. Zwężenie naczyń krwionośnych powoduje zmniejszenie dopływu krwi do skóry, co prowadzi do blednięcia skóry i zatrzymywania ciepła ciała. Ma to wpływ na reakcję walki lub ucieczki, zmniejszając przepływ krwi do skóry twarzy i przekierowując ją do ważniejszych obszarów, takich jak naczynia krwionośne mięśni.
System przedsionkowy
SCG jest połączona ze strukturami przedsionkowymi, w tym z nabłonkiem nerwowym kanałów półkolistych i narządami otolitowymi, stanowiąc możliwy substrat do modulacji odruchów przedsionkowo-współczulnych.
Znaczenie kliniczne
Zespół Hornera
Zespół Hornera to zaburzenie wynikające z uszkodzenia współczulnej autonomicznej ścieżki nerwowej w głowie. Uszkodzenie SCG, będącej częścią tego systemu, często prowadzi do zespołu Hornera. Uszkodzenie obszarów rdzenia kręgowego T1-T3 jest odpowiedzialne za opadanie powiek ( opadanie powiek ), zwężenie źrenicy ( mioza ) i zapadanie się gałki ocznej (pozorny Enophthalmos ; powieka). Uszkodzenie lub znaczne uszkodzenie SCG skutkuje zaburzeniem neuronów trzeciego rzędu (patrz Zespół Hornera: Patofizjologia ).
Rodzinna dysautonomia
Dysautonomia rodzinna to zaburzenie genetyczne charakteryzujące się nieprawidłowościami neuronów czuciowych i współczulnych. SCG jest znacząco dotknięty tą utratą neuronów i może być odpowiedzialny za niektóre z wynikających z tego objawów. W badaniach pośmiertnych SCG ma średnio jedną trzecią normalnej wielkości i ma tylko 12% normalnej liczby neuronów. Defekty w kodowaniu genetycznym NGF, które powodują mniej funkcjonalny NGF o nieprawidłowej strukturze, mogą być molekularną przyczyną dysautonomii rodzinnej. NGF jest niezbędny do przeżycia niektórych neuronów, więc utrata funkcji NGF może być przyczyną śmierci neuronów w SCG.
Historia
Ponowne unerwienie
Pod koniec XIX wieku John Langley odkrył, że górny zwój szyjny jest zorganizowany topograficznie. Po stymulacji niektórych obszarów zwoju szyjnego górnego pojawiał się odruch w określonych obszarach głowy. Jego odkrycia wykazały, że neurony przedzwojowe unerwiają określone neurony postganglionowe. W swoich dalszych badaniach nad zwojem szyjnym górnym Langley odkrył, że zwój szyjny górny ma charakter regeneracyjny. Langley odciął SCG powyżej części T1, powodując utratę odruchów. Pozostawione same sobie, włókna ponownie unerwiły SCG i przywrócone zostały początkowe odruchy autonomiczne, chociaż odzyskiwanie funkcji szyszynki było ograniczone. Kiedy Langley zerwał połączenia między SCG a obszarem T1–T5 rdzenia kręgowego i zastąpił SCG innym, SCG wciąż był unerwiony w tej samej części rdzenia kręgowego, co wcześniej. Kiedy zastąpił SCG zwojem T5, zwój miał tendencję do unerwienia przez tylną część rdzenia kręgowego (T4–T8). Zastąpienie oryginalnego SCG innym lub zwojem T5 poparło teorię Langley'a dotyczącą specyficzności topograficznej SCG.
Badania
Zwoje obwodowego autonomicznego układu nerwowego są powszechnie wykorzystywane do badania połączeń synaptycznych. Zwoje te są badane, ponieważ połączenia synaptyczne wykazują wiele podobieństw do ośrodkowego układu nerwowego (OUN) i są również stosunkowo dostępne. Łatwiej je badać niż OUN, ponieważ mają zdolność odrastania, której neurony w OUN nie mają. SCG jest często stosowany w tych badaniach jako jeden z większych zwojów. Obecnie neuronaukowcy badają takie zagadnienia dotyczące SCG, jak przeżycie i odrost neurytów neuronów SCG, neuroendokrynne aspekty SCG oraz struktura i szlaki SCG. Badania te są zwykle przeprowadzane na szczurach, świnkach morskich i królikach.
Składki historyczne
- E. Rubin badał rozwój SCG u płodów szczurów. Badania nad rozwojem nerwów w SCG mają wpływ na ogólny rozwój układu nerwowego.
- Wpływ wieku na dendrytyczną arboryzację neuronów współczulnych badano w SCG szczurów. Wyniki wykazały, że w SCG młodych szczurów występuje znaczny wzrost dendrytyczny, ale nie występuje u szczurów starszych. U starszych szczurów stwierdzono zmniejszenie liczby dendrytów.
- Komórki SCG wykorzystano do badania czynnika wzrostu nerwów (NGF) i jego zdolności do kierowania wzrostem neuronów. Wyniki pokazały, że NGF miał ten kierunkowy lub tropikalny wpływ na neurony, kierując kierunek ich wzrostu.
Dodatkowe obrazy

Prawy łańcuch współczulny i jego połączenia ze splotami piersiowymi, brzusznymi i miednicznymi.

Superior zwoju szyjnego

Połączenia współczulne zwojów rzęskowych i szyjnych górnych.
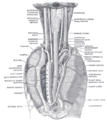
Położenie i relacja przełyku w odcinku szyjnym i śródpiersiu tylnym. Widziane z tyłu.

Unerwienie współczulnego tułowia i SCG narządów docelowych w głowie.





Bibliografia
![]() Ten artykuł zawiera tekst w domenie publicznej ze strony 978 20. wydania Anatomii Graya (1918)
Ten artykuł zawiera tekst w domenie publicznej ze strony 978 20. wydania Anatomii Graya (1918)
- ^ B Michael J. Zigmond, wyd. (2000). Podstawowa neuronauka (2 wyd.). San Diego: Acad. Naciskać. s. 1028–1032 . Numer ISBN 0127808701.
- ^ Henryk Szary. Anatomia ciała ludzkiego . 20. ed. Filadelfia: Lea i Febiger, 1918 Nowy Jork: Bartleby.com, 2000. http://www.bartleby.com/107/210.html . Dostęp 9 lipca 2013 r.
- ^ B Li Chen; Róg, John P. (2005). „Fizjologiczna klasyfikacja neuronów współczulnych w górnym zwoju szyjnym szczura”. Czasopismo Neurofizjologii . 95 (1): 187-195. doi : 10.1152/jn.00779.2005 . PMID 16177176 .
- ^ Purves, D; Wigston, DJ (styczeń 1983). „Jednostki nerwowe w górnym zwoju szyjnym świnki morskiej” . Czasopismo Fizjologii . 334 (1): 169–78. doi : 10.1113/jphysiol.1983.sp014487 . PMC 1197307 . PMID 6864556 .
- ^ Purnyn, H..; Rikhalsky, O.; Fedułowa S.; Veslovsky, N. (2007). „Ścieżki transmisji w górnym zwoju szyjnym szczura”. Neurofizjologia . 39 (4-5): 396-399. doi : 10.1007/s11062-007-0053-2 . S2CID 27184650 .
- ^ A B Purves Dale; Lichtman, Jeff W. (2000). Rozwój układu nerwowego . Sunderland, Massachusetts: Sinauer Associates. s. 236–238 . Numer ISBN 0878937447.
- ^ B Purves Dale (2012). Neuronauka (5 wyd.). Sunderland, Mass.: Sinauer. P. 465. ISBN 9780878936953.
- ^ Fotoperiodyzm, melatonina i szyszynka . Londyn: Pitman Publishing Ltd. 2009. s. 14.
- ^ Lichtman, Jeff W.; Purves, Dale; Yip, Joseph W. (1979). „W celu selektywnego unerwienia górnych komórek zwoju szyjnego świnki morskiej” . Czasopismo Fizjologii . 292 (1): 69–84. doi : 10.1113/jphysiol.1979.sp012839 . PMC 1280846 . PMID 490406 .
- ^ Pearson, J; Brandeis, L; Goldstein, M (5 października 1979). „Immunoreaktywność hydroksylazy tyrozynowej w rodzinnej dysautonomii”. Nauka . 206 (4414): 71-72. Kod Bibcode : 1979Sci...206...71P . doi : 10.1126/science.39339 . PMID 39339 .
- ^ Schwartz, JP; Breakefield, XO (luty 1980). „Zmieniony czynnik wzrostu nerwów w fibroblastach od pacjentów z rodzinną dysautonomią” . Materiały Narodowej Akademii Nauk Stanów Zjednoczonych Ameryki . 77 (2): 1154–8. Kod Bib : 1980PNAS...77.1154S . doi : 10.1073/pnas.77.2.1154 . PMC 348443 . PMID 6244581 .
- ^ Sanes, Dan H.; Reh, Tomasz A.; Harris, William A. (1985). Zasady rozwoju neuronowego . San Diego, Kalifornia: Prasa akademicka. s. 214-221. Numer ISBN 0-12-300330-X.
- ^ Lingappa, Jaisri R .; Zigmond, Richard E. (2013). „Ograniczone odzyskiwanie funkcji szyszynki po regeneracji przedzwojowych aksonów współczulnych: dowody na utratę zwojowej specyficzności synaptycznej” . Dziennik Neuronauki . 33 (11): 4867–4874. doi : 10.1523/JNEUROSCI.3829-12.2013 . PMC 3640627 . PMID 23486957 .
- ^ Purves, D; Lichtman, JW (październik 1978). „Tworzenie i utrzymanie połączeń synaptycznych w zwojach autonomicznych”. Recenzje fizjologiczne . 58 (4): 821–62. doi : 10.1152/physrev.1978.58.4.821 . PMID 360252 .
- ^ Rubin, E (marzec 1985). „Rozwój zwoju szyjnego górnego szczura: dojrzewanie komórek zwojowych” . Dziennik Neuronauki . 5 (3): 673–84. doi : 10.1523/jneurosci.05-03-00673.1985 . PMC 6565020 . PMID 2983044 .
- ^ Andrews, TJ; Li, D; Halliwell, J; Cowen, T (luty 1994). „Wpływ wieku na dendryty w zwoju szyjnym górnym szczura” . Dziennik Anatomii . 184 (1): 111–7. PMC 1259932 . PMID 8157483 .
- ^ Campenot, RB (1977). „Lokalna kontrola rozwoju neurytów przez czynnik wzrostu nerwów” . Proc Natl Acad Sci USA . 74 (10): 4516–9. Kod Bibcode : 1977PNAS...74.4516C . doi : 10.1073/pnas.74.10.4516 . PMC 431975 . PMID 270699 .
Zewnętrzne linki
- Zdjęcie anatomiczne: 31:07-0201 w SUNY Downstate Medical Center